Front Immunol:靶向NKG2D配体激活NK细胞
时间:2023-09-02 13:59:26 热度:37.1℃ 作者:网络
免疫检查点抑制剂(ICI)彻底改变了免疫肿瘤学领域,部分患者可以实现肿瘤的持久免疫控制。然而,肿瘤免疫逃避机制依然存在,如何让更多的病人获益,开发更有效的治疗方案始终面临着挑战。
目前,联合疗法正成为免疫肿瘤学研究的中心。这种联合疗法应该增强抗肿瘤免疫反应或靶向肿瘤免疫逃逸机制,特别是肿瘤微环境(TME)中的各种参与者。自然杀伤(NK)细胞最近处于许多免疫治疗策略的前沿,人们正在开发一些新的方法来充分利用NK细胞的抗肿瘤潜力。
最相关的NK细胞激活受体之一是NKG2D,其识别8种不同的NKG2D配体(NKG2DL),包括MICA和MICB。MICA和MICB在正常细胞上低表达,但在受损、转化或感染的细胞表面表达上调,它们与NKG2D的结合触发NK细胞效应器功能。此外,MICA/B是多态性的,这种多态性通过调节其细胞表面表达、细胞内运输、免疫抑制的可溶性形式的脱落或NKG2D相互作用的亲和力来影响功能反应。
目前,针对NKG2D-NKG2DL轴的免疫治疗方法正在研究中,通过重塑TME,释放NK细胞和细胞毒性CD8+T细胞的抗肿瘤效应,为更多的肿瘤病人获益带来希望。
NK细胞在肿瘤免疫中起着关键作用,目前,NK细胞逐渐成为当前免疫治疗策略的前沿领域。许多新化合物包括单克隆抗体正在开发中,以充分发挥其抗肿瘤潜力。此外,NK细胞的过继疗法包括异基因NK细胞和CAR-NK,这些基于NK细胞的策略已经取得了一些成功。
CAR-NK细胞比CAR-T细胞有几个明显优势,CAR-NK 细胞的寿命很短,几乎不会产生靶外效应。另外,NK细胞产生的细胞因子种类与T淋巴细胞产生的细胞因子种类有很大不同。活跃的NK细胞通常产生IFN-γ和粒-巨噬细胞集落刺激因子(GM-CSF),不会产生CAR-T细胞通常诱导的细胞因子风暴。
然而,实体瘤的情况大不相同,这主要是因为NK细胞必须面对克服免疫抑制性TME的艰巨任务,以避免耗竭和功能失调。此外,即使NK细胞能够克服这种不利环境,它们浸润实体瘤的能力较弱也是影响其疗效的一大原因。因此,NK细胞过继转移可能需要结合其他策略来增强NK细胞有效的抗肿瘤功能。与ICI的结合呈现出诱人的可能性。
临床中的单细胞RNA测序(scRNAseq)数据表明,肿瘤NK细胞浸润与几种不同癌症类型的更好预后相关,并且NK细胞浸润有助于产生强烈的ICI反应。此外,scRNAseq和CYTOF显示ICI诱导TME中的淋巴和髓系细胞的显著重塑,并且这种效应依赖于IFN-γ。因此,通过开发新的肿瘤免疫疗法来利用抗肿瘤NK细胞效应器功能引起了人们极大的兴趣。
NK细胞表面表达激活性受体和抑制性受体,当激活受体触发的信号超出抑制受体触发的信号时,NK细胞产生效应器功能。NK细胞活性也受细胞因子调节,尤其是由髓系细胞产生的细胞因子,如IL-12、IL-23、IL-27、IL-15、IL-18和TGF-β,以及由NK细胞表达的几种Toll样受体(如TLR3、TLR7和TLR9)激动剂。因此,环境中激活和抑制信号的整合极大地决定了NK细胞动员效应器功能的能力。
除了识别几个IgG亚类的Fc部分负责ADCC活性的CD16(FcγIIIa受体)外,自然细胞毒性受体(NCR)包括NKp30(CD337)、NKp44(CD336)、NKp46(CD335)、NKp80以及DNAM-1(CD226)和NKG2D(CD314)是主要的激活性受体。在动物模型中,阻断NKG2D或基因敲除可导致对肿瘤发生和进展的易感性增加。因此,人们正在努力利用NKG2D配体作为I-O的分子靶点。
在人类中,已经描述了8种不同的NKG2DL。第一个已知的NKG2DL是由MHC I类链相关基因A和B(MICA和MICB)编码的蛋白质,也分别称为PERB11.1和PERB11.2。这两个基因都位于MHC内,具有高度多态性,并且等位基因以共显性方式表达。大多数等位基因编码的MICA和MICB蛋白由三个胞外结构域、一个跨膜结构域和一个胞内尾部组成。
其余的6个NKG2DL是UL-16结合蛋白(ULBP)家族的成员,也称为维甲酸早期转录物(RAET)1。它们分别被命名为ULBP-1(RAET1I)、ULBP-2(RAET1H)、ULBP-3(RAET1N)、ULBP-4(RAET1E)、ULBP-5(RAET1G)和ULBP-6(RAET1L)。
MICA在多种肿瘤中均有表达,RNA测序数据表明,MICA在肺癌、结直肠癌、胃癌、肝癌和乳腺癌中表达最高。此外,MICA显示出非常低的肿瘤突变负担,这表明其表达不受DNA编辑的影响,从而赋予肿瘤某种适应性优势。尽管NKG2DL的过度表达可能是限制肿瘤进展的有效策略,但肿瘤同时也显示出破坏NKG2DL生物学功能的逃逸策略,包括由肿瘤分泌的金属蛋白酶(MMP)诱导分泌的可溶性MICA。可溶性MICA(sMICA)和可溶性MICB(sMICB)随后可与NKG2D结合并诱导其下调和降解,破坏NK细胞的效应器功能并促进肿瘤免疫逃逸。

鉴NK细胞可以作为肿瘤炎症的驱动因素,导致肿瘤免疫细胞浸润,并促进“冷”肿瘤转化为“热”肿瘤,从而提高对ICI的反应性。NK细胞诱导有效肿瘤免疫的关键功能取决于与传统树突状细胞(cDC1)的成功串扰以及趋化因子CCL5和XCL1的产生,并且肿瘤源性前列腺素E2(PGE2)干扰这种相互刺激。
此外,TGF-β是NK细胞效应器功能的主要负调节因子。体外试验显示,TGF-β和MMP产生的自分泌回路促进MICA和ULBP-2的脱落,并对肿瘤细胞表面上这些NKG2DL的表达产生负面影响。TGF-β还抑制NK细胞上NKp30和NKG2D的表达。TGF-β还影响NK细胞代谢,导致糖酵解和氧化磷酸化减少,从而抑制NK细胞效应器功能。因此,干扰TGF-β的新疗法可能触发NKG2D依赖性NK细胞介导的肿瘤消除。
另一个导致NK细胞功能障碍的因素是慢性刺激。过表达MICA的小鼠触发慢性刺激导致NK细胞NKG2D表达减少,这些NK细胞表现出受损的NKG2D依赖的细胞毒性。
TAM也是肿瘤免疫逃逸的参与者。靶向TAM可以通过恢复NK细胞活化和效应器功能来改善肿瘤免疫,通过操纵TAM和NK细胞将“冷”肿瘤转化为“热”肿瘤的策略可能构成I-O的前沿策略。
抑制性受体构成的免疫检查点以PD-1/PD-L1轴为代表,包括CD96、NKG2A、TIGIT、TIM-3和LAG-3已成为最前沿的研究热点,而它们也通常在功能失调的NK细胞中过度表达。
越来越多的证据表明NK细胞也表达PD-1和PD-L1,并且以它们为靶点,可以恢复抗肿瘤NK细胞的效应器功能。在几种胃肠道肿瘤患者的循环NK细胞(PBNK)和肿瘤浸润NK(TINK)细胞中观察到PD-1+NK细胞频率增加,PD-1表达增强,并且在某些情况下,PD-1的高表达与生存受损相关。
针对NK细胞上的其他共抑制受体也在研究中。TIGIT是一种共抑制受体,它在耗竭的TINK和肿瘤浸润性T细胞上过度表达。阻断TIGIT可诱导NK细胞恢复活力,恢复有效的肿瘤免疫,并提高PD-1抑制剂的疗效。
CD96是NK细胞上表达的另一种共抑制受体,通过与肿瘤细胞上表达的CD155结合,限制NK细胞效应器功能。在无病生存率降低的肝细胞癌患者中,出现TINK耗竭,CD96+细胞频率升高,CD96表达增加,但阻断CD96可恢复NK细胞介导的效应器功能。
NKG2A是另一种与CD94相关的抑制性受体,NKG2A及其配体HLA-E的高表达可在肝细胞癌患者的肿瘤组织中检测到,表达NKG2A的TINK表现出耗竭细胞的特征,并与不良预后相关。NKG2A和HLA-E在其他几种人类癌症中过度表达,用Monalizumab(一种人源化抗NKG2A单抗)阻断NKG2A,通过促进NK细胞和CTL细胞效应器功能,与抗PD-L1的ICI联合应用增强肿瘤免疫。
此外,慢性淋巴细胞白血病(CLL)患者在白血病母细胞、NK细胞和T淋巴细胞上表现出LAG-3上调,伴有血浆中大量可溶性LAG-3,与预后受损相关。使用relatlimab(一种抗LAG-3单克隆抗体),可以清除白血病细胞并恢复NK细胞和T细胞效应器功能。相反,抑制性受体KIR2DL1、KIR2DL2和KIR2DL3的阻断性单克隆抗体Lirilumab并未显示预期的临床疗效,这表明需要仔细选择靶向共抑制受体。
MICA是肿瘤中广泛表达的NKG2DL,其作为IO策略的靶点具有应用前景。ADCC是NK细胞介导的主要效应器功能之一, 利用NK细胞介导的ADCC作用开发靶向MICA的单克隆抗体是主要策略之一。
此外,在ipilimumab治疗的黑色素瘤患者中可以观察到自体产生的抗MICA抗体,可促进从血浆中清除sMICA,并促进树突状细胞对肿瘤抗原的交叉呈递。

BLS-MICA是一种MICA胞外结构域与具有佐剂特性的细菌免疫原性蛋白融合而成的嵌合蛋白。BLS-MICA在体内可以诱导高滴度的抗MICA抗体,这种“二合一”策略,一方面通过ADCC作用消除肿瘤,另一方面通过清除sMICA干扰肿瘤免疫逃逸。
此外,一些抗MIC的单克隆抗体也显示出有希望的临床前结果。靶向sMICA(B10G5)的单克隆抗体单独或与抗CTLA-4单克隆抗体联合应用显示出治疗效果。其他针对α3结构域的抗MICA/B单克隆抗体抑制了蛋白水解脱落。其中一种单抗(7C6)抑制MICA和MICB脱落,延缓了小鼠黑色素瘤和结肠癌的生长。此外,7C6通过稳定肿瘤细胞表面MICA/B表达与HDACi panobinostat和CIK产生协同作用。
利用CAR-T或CAR-NK也可以靶向NKG2DL。已开发出一些经工程设计表达NKG2D的CAR-T细胞,其在体外对细胞系和体内对异种移植的人或同基因小鼠肿瘤细胞系具有增强的效应器功能。此外,表达人NKG2D胞外结构域与DAP12融合的CAR-NK细胞在体外对几种实体瘤细胞系表现出增强的NK细胞介导的细胞毒性,并在体内显示出治疗效果。在三名结直肠癌患者中,这些CAR-NK细胞介导肿瘤消退和患者临床改善,突出了它们在实体瘤中的潜在治疗效用。
目前使用ICI的联合疗法无疑是一种趋势,通过与其他药物联合来充分发挥抗MICA/B抗体的治疗效用。以下的一些药物可能是与抗MICA/B抗体组合的有希望的候选者。
促进肿瘤细胞MICA/B上调的药物
NKG2DL的表达受DNA损伤反应途径(DDR)的控制,但是组蛋白脱乙酰酶抑制剂(HDACi)等表观遗传调节剂也可以触发MICA/B的上调表达。HDACi通过诱导细胞周期阻滞、凋亡和自噬发挥抗增殖作用,已观察到TSA、SAHA、belinostat、VPA、NaB、romidepsin和Nosentat在来源于液体和实体肿瘤的不同细胞系中触发MICA/B的上调。
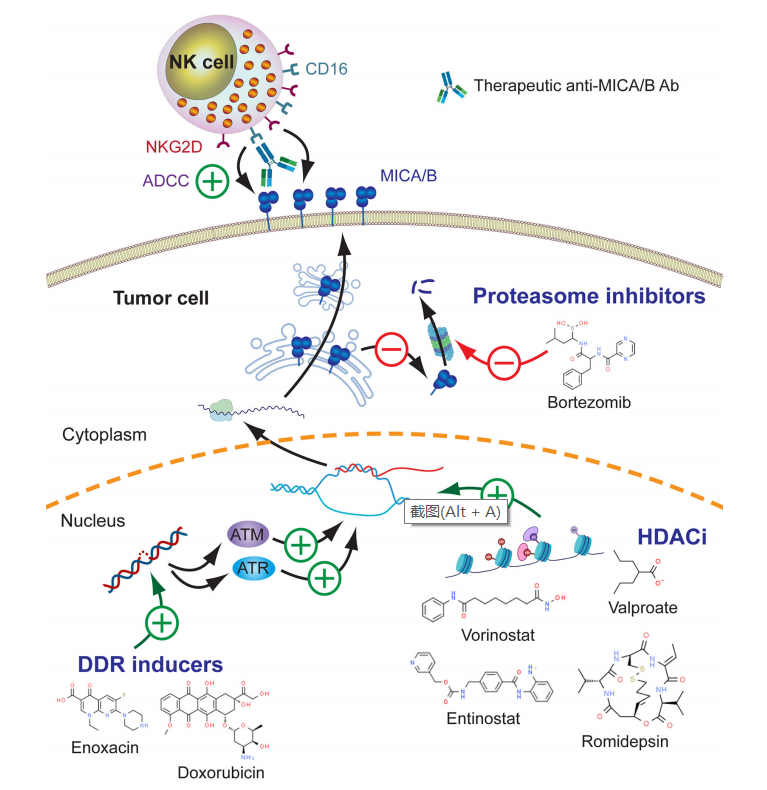
综合关于HDACi对MICA/B表达、NKG2D表达和NK细胞效应器功能影响的数据,已获批的少数HDACi,如Vorinotsat,belinostat和entinostat可能成为与抗MICA/B抗体产生协同作用的敏感分子。
Bortezomib(一种用于治疗MM患者的蛋白酶体抑制剂)也能上调MICA/B的表达。另外用于治疗癌症患者的其他药物,如索拉非尼和舒尼替尼以及5-氟尿嘧啶也可以影响MICA/B的表达。总之,目前用于治疗不同类型癌症患者的几种药物可能刺激MICA/B的上调,这可能增加肿瘤细胞对抗MICA/B抗体和NK细胞的敏感性。
MICA/B脱落阻断剂
MICA/B的脱落取决于几种MMP介导的蛋白水解裂解,如ADAM10、ADAM17和MMP14或胞外体的分泌。因此,阻断MICA/B的脱落将干扰由这些可溶形式的MICA/B驱动的肿瘤免疫逃逸,也可以回复MICA/B的细胞表面表达和NKG2D依赖性NK细胞效应器功能的改善。因此,MMP抑制剂(MMPI)是极具吸引力的候选药物,如MMPI-I、MMPI-II、MMPI-III、MMPI-IV、batimastat、BB94、GW280264X、GI1254023X、ilomastat、 GM6001、URB597、periostat和一些ADAM-10选择性抑制剂。
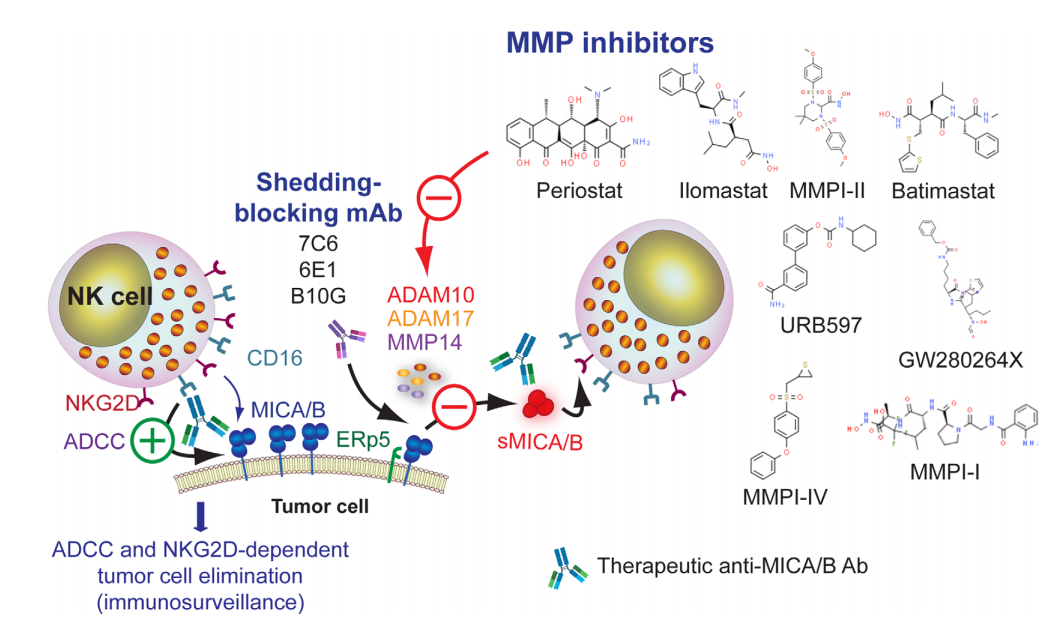
然而,在这些化合物中,目前只有periostat用于不同类型肿瘤的临床试验。此外,单克隆抗体7C6和6E1都识别MICA/B的α3结构域,抑制其蛋白水解脱落,并导致MICA/B的细胞表面表达增加。总之,对MICA/B脱落的抑制构成了与抗MICA/B抗体结合以诱导改善ADCC和肿瘤细胞清除的另一个机会。
PARP抑制剂
合成致死性诱导剂,如olaparib、rucaparib、niraparib、talazoparib、veliparib等,由于其抑制PARP1(DNA修复的关键酶)的能力而发挥治疗作用。PARP1抑制也被证明影响NKG2D/NKG2DL轴,观察发现PARP1参与AML患者白血病干细胞(LSC)中NKG2DL(主要是MICA和MICB)的抑制。

抑制PARP1会释放AML细胞上NKG2DL的表达,进而使其更易受NKG2D依赖性、NK细胞介导的效应器功能的影响,肿瘤细胞得到有效的清除。此外,PARP抑制剂和CSF-1R阻断剂的组合增强了体内BRCA缺陷肿瘤的抗肿瘤免疫并延长生存期,表明PARP抑制影响TME中的TAM抑制活性。总的来说,这些结果为PARP1抑制剂和靶向MICA/B的抗体之间的联合疗法提供了坚实的理论基础。
STING激动剂
PARP1抑制的下游效应涉及cGAS-STING途径的激活,STING通过产生I型IFN增强NK细胞激活和肿瘤免疫。因此,STING激动剂可将TME重新配置为促炎症环境,并促进“冷”肿瘤转化为“热”肿瘤。STING激活还与ICI联合,通过TAM重编程诱导抗肿瘤免疫。此外,STING激动剂可以刺激NK细胞介导的CD8+T细胞耐药肿瘤的清除,并动员肿瘤浸润的髓系细胞产生IFN-β,进而激活NK细胞。
由于cGAS/STING途径的激活诱导NKG2DL的上调,STING已成为IO中另一个有吸引力的分子靶点,以利用NKG2D依赖性NK细胞介导的抗肿瘤效应,与抗MICA/B的抗体联合从而调节TME并诱导抗肿瘤免疫。
诱导免疫原性细胞死亡的药物
诱导ICD的药物,如蒽环类药物可以触发有效的抗肿瘤免疫反应,抑制小鼠肿瘤生长,因为它们使肿瘤细胞具有免疫原性。这一过程涉及钙网蛋白(CRT)向肿瘤细胞表面的动员,ATP和几种警报蛋白(如HMGB1和膜联蛋白A1)的分泌和细胞外积累,以及I型IFN的产生,它们共同作用,促进重塑TME的炎症反应,抑制TAM并对肿瘤生长产生负面影响。

目前许多临床试验正在探索触发ICD的化合物,单药或与ICI联合,通过刺激死亡肿瘤细胞的免疫原性来促进肿瘤免疫。ICD诱导剂也可以与抗MICA/B抗体联合,以增强针对NK细胞介导的ADCC,并通过涉及TME重塑和TAM重编程的效应以防止耗竭。
靶向TAM的分子
TAM的重编程可以使其转换为促炎巨噬细胞,在体外和体内恢复NK细胞效应器功能。Tyro、Axl和MerTK受体酪氨酸激酶(TAMRTK)是免疫反应内稳态的重要参与者,因为它们参与炎症的解决,有助于清除凋亡细胞碎片,恢复血管完整性,并调节免疫反应的幅度。因此,干扰TAMRTK功能可导致慢性炎症和自身免疫。

TAMRTK的表达和信号转导的改变与肿瘤进展有关,阻断TAM上的TAMRTK可通过触发STING激活,刺激I型IFN反应,重塑TME,促进T细胞活化,并与ICI协同促进有效的抗肿瘤免疫。目前,人们正在努力开发TAMRTK抑制剂。
NK细胞目前是肿瘤免疫治疗领域的重要组成部分,具有极大的治疗潜力。在血液肿瘤的治疗已经取得了一些初步的成功,但实体瘤的治疗依然存在很多难题,尤其是要克服免疫抑制性的TME。
目前,选择新的靶分子和治疗方式是一个重要方向。靶向NKG2D/NKG2DL轴,尤其是其中的MICA和MICB是一个非常有吸引力的靶点,利用NKG2D依赖性NK细胞介导的抗肿瘤作用促进肿瘤免疫。抗MICA/B抗体可以和多种药物联合,所有这些策略都直接或间接地利用了MICA/B,通过恢复NK细胞激活和效应器功能来消除肿瘤,并将“冷”肿瘤转化为“热”肿瘤。
未来还可能有一些新的策略,如针对MICA/B的基于CAR-T/CAR-NK细胞的治疗,溶瘤病毒,基因编辑等创新的免疫治疗模式,惠及更广泛的癌症患者。
参考文献:
1.Leveraging NKG2D Ligands inImmuno-Oncology. Front Immunol. 2021; 12: 713158.

